『BCNEセンターについての研究』
(序章)
 1924年、ドイツの科学者であるハンス・シュペーマン教授とヒルデ・マンゴールド研究員は、両生類胚の原口上唇部領域に非常に強い誘導能を持つ領域が存在することを発見した。右図は、その領域を二つもつ胚を黒田がアフリカツメガエルを用いて作成したものであるが、頭部から尾部構造までが完全に保存された二次軸胚が原口上唇部によって誘導されている(このような技術を胚操作と呼ぶ:黒田は世界でもトップ10に入る胚操作技術を持つと自負している)。この偉大な発見によりシュペーマンは1935年にノーベル医学生理学賞を受賞し、また、この偉大な領域はシュペーマン・オーガナイザーと呼ばれるようになった。 1924年、ドイツの科学者であるハンス・シュペーマン教授とヒルデ・マンゴールド研究員は、両生類胚の原口上唇部領域に非常に強い誘導能を持つ領域が存在することを発見した。右図は、その領域を二つもつ胚を黒田がアフリカツメガエルを用いて作成したものであるが、頭部から尾部構造までが完全に保存された二次軸胚が原口上唇部によって誘導されている(このような技術を胚操作と呼ぶ:黒田は世界でもトップ10に入る胚操作技術を持つと自負している)。この偉大な発見によりシュペーマンは1935年にノーベル医学生理学賞を受賞し、また、この偉大な領域はシュペーマン・オーガナイザーと呼ばれるようになった。
この発見は、生物学の世界に非常に大きな影響をもたらし、発生生物学者の殆どが中胚葉であるシュペーマンオーガナイザーを中心として全ての組織が誘導されていくと考えるようになった。シュペーマン・オーガナイザーを発生の総司令官として捉える考え方だ。
しかし、20世紀末になり、シュペーマン・オーガナイザーが形成される時期に発現する遺伝子群の詳細な分子情報が明らかになるにつれ、神経(特に脳などの前方神経に関して)はこの考え方に当てはまらない結果が出始めた。なぜなら、中胚葉が全くできない条件下(つまりシュペーマン・オーガナイザーが存在できない条件下)において、依然として前方の神経が誘導されることが両生類や魚類を用いた実験において確認されたからである。つまり、総司令官がいなくても脳組織が作られるという訳だ。また、外胚葉領域に発現するBMP (Bone Morphogenetic Protein)と呼ばれる分泌タンパク質の活性が阻害されることによって、外胚葉領域は中胚葉とは無関係に神経化することも発見された。21世紀になり、シュペーマン・オーガナイザー絶対論は崩れ始めたわけである。
ちなみに、発生学者の大御所の先生の中には「シュペーマン・オーガナイザーの発見が発生学を四半世紀遅らせた」とおっしゃる方もいます。私はそこまで極端な意見は有していませんが、盲目的なまでのシュペーマン・オーガナイザー信奉がいくつかの真実にフィルターをかけてしまったことは間違いないと思います。
(私のこれまでの研究)
なぜ、中胚葉が存在しないにも関わらず神経が誘導されるのであろうか。その理由は、外胚葉領域において神経を誘導できる物質が発現しているからに違いないと考えるのが、最も素直な論理であろう。
 私はまず、上に述べた中胚葉が存在しない胚を人工的に作成した時に、胞胚期において背側動物極側の領域(右に示す模式図の緑色部分)にBMPの働きを阻害するタンパク質(BMPアンタゴニストとも呼べる)であるChordinとNogginが発現していることに気づいた。しかし、だからといって、それが実際に神経を誘導する原因となっているとは限らない。そこで、中胚葉不在の胚において神経が形成される領域がその領域に由来するかどうかを確かめた。その結果、神経に分化した領域は100%その領域に由来していることを突き止めた。また、BMPアンタゴニストを中胚葉非存在下の胚において働かないようにすることによって、神経形成が阻害されることも発見した。これらの理由より、原腸胚のシュペーマン・オーガナイザーの出現よりも前にあたる胞胚期において発現するChordinとNogginの存在が、中胚葉非存在下の胚における神経誘導に関わっていることが証明された訳である。それ故、この特殊な領域を、胞胚期(blastula)においてChordinとNogginが発現する領域として、BCNEセンター(Blastula Chordin- and Noggin-Expressing center)と名づけた。 私はまず、上に述べた中胚葉が存在しない胚を人工的に作成した時に、胞胚期において背側動物極側の領域(右に示す模式図の緑色部分)にBMPの働きを阻害するタンパク質(BMPアンタゴニストとも呼べる)であるChordinとNogginが発現していることに気づいた。しかし、だからといって、それが実際に神経を誘導する原因となっているとは限らない。そこで、中胚葉不在の胚において神経が形成される領域がその領域に由来するかどうかを確かめた。その結果、神経に分化した領域は100%その領域に由来していることを突き止めた。また、BMPアンタゴニストを中胚葉非存在下の胚において働かないようにすることによって、神経形成が阻害されることも発見した。これらの理由より、原腸胚のシュペーマン・オーガナイザーの出現よりも前にあたる胞胚期において発現するChordinとNogginの存在が、中胚葉非存在下の胚における神経誘導に関わっていることが証明された訳である。それ故、この特殊な領域を、胞胚期(blastula)においてChordinとNogginが発現する領域として、BCNEセンター(Blastula Chordin- and Noggin-Expressing center)と名づけた。
では、シュペーマン・オーガナイザーが存在する胚(正常胚)において、BCNEセンターは脳領域に分化しているのであろうか。この疑問に答えるために、私はBCNEセンターの詳細な運命追跡実験を行った。その結果、正常胚においても脳の殆どの領域がBCNEセンターに依存していることが判明した(右図のオタマジャクシの緑色領域がBCNEセンターに由来している)。つまり、両生類胚はシュペーマン・オーガナイザーが出現する前から、脳ができる領域を予め準備していたことになる。この発見は、発生生物学の中で謎とされていたシュペーマンオーガナイザーと接していない外胚葉領域において起こる神経誘導(側方神経誘導現象)に関しても、明確な答えを与えることになった。
この研究内容は、世界最高峰の科学雑誌であるNature誌からも高く評価され、神経発生の企画において、マイルストーンにあたる論文として紹介される栄誉を得た。
(現在のBCNEセンターに関する研究)
BCNEセンターの役割に関する研究はこれで終わったわけではない。私達のグループは、BCNEセンターの半分近くの領域は脳以外の組織に分化することを確かめているからだ。実は、非常に面白いことに、シュペーマン・オーガナイザー領域は全てBCNEセンターに由来していることが判明している。つまり、原腸胚のシュペーマン・オーガナイザーは元々はBCNEセンターであったのだ。そう、BCNEセンターであったことが、将来、シュペーマン・オーガナイザーになるための非常に重要な条件であると考えたとして何の不思議があろうか。私達は現在、BCNEセンターの存在が、シュペーマン・オーガナイザーの形成において、どのような存在であるのかを検証する実験に取り組んでいる。この研究を進めることは、試験管内において効果的にシュペーマン・オーガナイザーを創り出すことにつながり、いずれは表皮領域だけで胚の全体構造を誘導させ、泳ぐオタマジャクシの創出につながるのではないか、と期待している。
それは、ある意味、フランケンシュタインの創造に近い実験と言えよう。このような実験は哺乳類を用いて行うには、非常に倫理的に反感を買う内容である。いや、それ依然に哺乳類胚を用いて細やかな胚操作を行った胚を十分な例数を得ることに無理がある。アフリカツメガエルを用いた本研究の推進こそが、最も進むべき道であると言えるだろう。
|
|
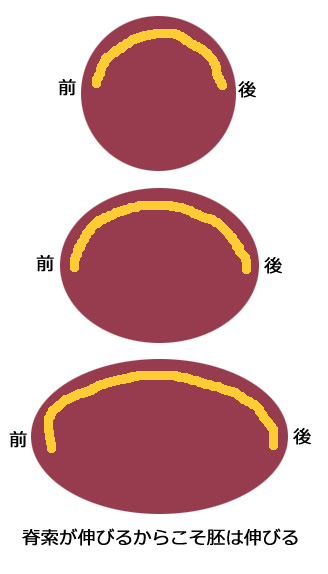 『脊索細胞の集合に関わる分子Axial protocadherin』
『脊索細胞の集合に関わる分子Axial protocadherin』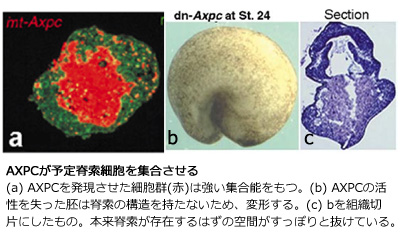 それでは、脊索を集合させる分子は何なのであろうか?予定脊索細胞がカルシウムの不在時にバラバラになること、そして、細胞同士が強い接着性を有することから、私はカドヘリンファミリーに属する分子がその役割を担っていることを考え、カドヘリンファミリーに従ったスクリーニングを行った。その結果、脊索特異的に発現する分子Axial protocadherin (AXPC)を得ることができた。このprotocadherinは一般的なcadherinファミリーと違い、1) 細胞外の繰り返しドメインが6つあり(一般的には5つ)、また細胞内にbeta-cateninの結合領域を有していなかった。gain-of-functionalな機能解析を行った結果、AXPCを発現させた外胚葉細胞は、脊索と同様の挙動を示し、強い集合能力を有することが分かった。また、loss-of-functionalな機能解析を行った結果、AXPCが働くことの出来ない脊索は、集合能力を欠失することが分かった。AXPCが働かなくなった胚では脊索領域がポッカリと無くなり、脊索細胞の分散によって生じた異所的な神経構造の存在が確認された。以上の結果より、脊索を組織として脊索たらしめていたものはAXPCであったことがわかる。このAXPCの存在なくしては脊椎動物は、その形に至るスタートを切ることも許されない訳である。
AXPCはヒトにおいても発見され、protocadherin-1という名称でも呼ばれている。また、腎臓において発現するなど、脊索以外での活躍も認められる。脊椎動物が、非常に強い細胞集合能を必要とした際にAXPCが使用され、その一番最初の使用用途が脊索の集合ということなのかもしれない(
それでは、脊索を集合させる分子は何なのであろうか?予定脊索細胞がカルシウムの不在時にバラバラになること、そして、細胞同士が強い接着性を有することから、私はカドヘリンファミリーに属する分子がその役割を担っていることを考え、カドヘリンファミリーに従ったスクリーニングを行った。その結果、脊索特異的に発現する分子Axial protocadherin (AXPC)を得ることができた。このprotocadherinは一般的なcadherinファミリーと違い、1) 細胞外の繰り返しドメインが6つあり(一般的には5つ)、また細胞内にbeta-cateninの結合領域を有していなかった。gain-of-functionalな機能解析を行った結果、AXPCを発現させた外胚葉細胞は、脊索と同様の挙動を示し、強い集合能力を有することが分かった。また、loss-of-functionalな機能解析を行った結果、AXPCが働くことの出来ない脊索は、集合能力を欠失することが分かった。AXPCが働かなくなった胚では脊索領域がポッカリと無くなり、脊索細胞の分散によって生じた異所的な神経構造の存在が確認された。以上の結果より、脊索を組織として脊索たらしめていたものはAXPCであったことがわかる。このAXPCの存在なくしては脊椎動物は、その形に至るスタートを切ることも許されない訳である。
AXPCはヒトにおいても発見され、protocadherin-1という名称でも呼ばれている。また、腎臓において発現するなど、脊索以外での活躍も認められる。脊椎動物が、非常に強い細胞集合能を必要とした際にAXPCが使用され、その一番最初の使用用途が脊索の集合ということなのかもしれない(